黑色素瘤的分子分類 — 第三部分 (Molecular Classification of Melanoma — Part 3)
日光暴露皮膚黑色素瘤的其他基因變異
在日光暴露皮膚 (sun-exposed skin) 的黑色素瘤 (melanoma) 中,已發現許多其他反覆出現的基因變異,包括 MITF 的擴增 (amplification) 或突變,以及編碼 β-catenin 的 CTNNB1 之活化突變 (activating mutation)。MITF 是一種 basic helix-loop-helix (hHLH)-leucine zipper 蛋白,在黑色素細胞 (melanocyte) 的發育中扮演角色。β-catenin 是 WNT pathway 的關鍵訊息傳遞組件,於其中作為一種黏著連接 (adherens junction) 蛋白,同時也能作為轉錄因子 (transcription factor),並在一小部分黑色素瘤中發生突變。此外,端粒相關蛋白 (telomere-related proteins) 為黑色素瘤發展的關鍵因子,這點也愈來愈明確。端粒 shelterin 複合體的組件(例如 POT1、ACD、TERF2IP)負責封蓋端粒末端並防止重組事件 (recombination events),其胚系突變 (germline mutations) 已與家族性黑色素瘤 (familial melanoma) 相關聯。TERT(編碼 telomerase 之蛋白組件的基因)啟動子 (promoter) 的突變,亦曾在家族性黑色素瘤以及百分之三十五至百分之七十的偶發性黑色素瘤 (sporadic melanomas) 中被描述。TERT promoter 突變是黑色素瘤中最常見的突變,並具有 UV 標記 (UV signature) 的 C > T 模式。TERT 與其他端粒相關因子的易感性突變 (predisposing mutations) 被認為會藉由增加端粒長度、延長腫瘤細胞的複製壽命 (replicative lifespan),進而增加獲得轉化 (transformation) 所需額外突變的可能性,從而提高黑色素瘤的風險。許多其他基因亦被認為與黑色素瘤有關,其中多數列於 Table 26.3。
紫外線受保護部位的黑色素瘤 (Melanomas on UV-protected sites)
肢端與黏膜黑色素瘤 (Acral and mucosal melanoma)
手部與足部的掌蹠表面 (volar surfaces) 極少暴露於 UV 輻射,且無毛皮膚 (glabrous skin) 具有較厚的表皮與極厚的角質層 (stratum)。
基因擴增 (gene amplifications) 典型上經由雙股 DNA 斷裂 (double-stranded DNA breaks) 與後續染色分體 (chromatids) 端對端融合的反覆循環而產生,因此意味著對基因組完整性 (genomic integrity) 之控制的喪失。此種控制喪失與後續基因組不穩定 (genetic instability) 的原因目前尚不清楚,因為 TP53 與其他已知會誘發不穩定的突變在這些黑色素瘤中並不常見。雖然顯然 UV 輻射在這些黑色素瘤中並不扮演重要角色,但目前仍未釐清在肢端與黏膜黑色素瘤 (acral and mucosal melanomas) 的致病機轉中是否有特定致癌物 (carcinogens) 參與,或它們是否在無外在影響下因隨機性 (stochastic) 變異而發生。日光受保護部位的黑色素瘤與日光暴露皮膚黑色素瘤之間,在基因組不穩定程度與突變負荷 (mutation burden) 上的顯著差異,顯示這些黑色素瘤亞型之間的基因損傷類型有根本上的不同。在日光暴露的黑色素瘤中,UV 光產物 (UV photoproducts) 與氧化損傷 (oxidative damage) 是腫瘤生成 (tumorigenesis) 的主要驅動因子,而雙股 DNA 斷裂(作為斷裂—融合—橋接循環 breakage-fusion-bridge cycles 的一部分)以及/或其他尚待
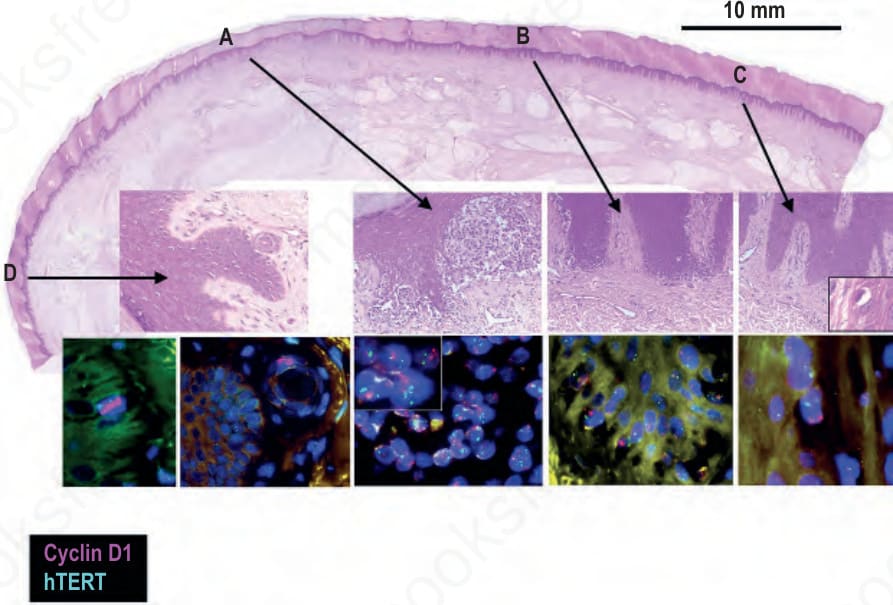
圖 26-139:肢端黑色素瘤中的場細胞 (field cells):上方面板顯示一個肢端黑色素瘤 (acral melanoma),其下方為四個特定區域的特寫。區域 A 顯示淺表侵襲性黑色素瘤 (superficially invasive melanoma),以 FISH 顯示 CCND1 (cyclin D1) 與 TERT (telomerase reverse transcriptase) 的擴增(分別為藍染細胞核中成簇的紅色與綠色訊號)。區域 B 顯示原位黑色素瘤 (melanoma in situ),具有 CCND1 擴增及正常的 hTERT 套數 (copy number)。區域 C 與 D 顯示組織病理學上未受侵犯的表皮,經 FISH 顯示帶有單一基底黑色素細胞具 CCND1 擴增(藍染細胞核內成簇的紅色訊號)。在區域 D 的第二個 FISH 面板中,一個具 CCND1 擴增的黑色素細胞位於一個 eccrine duct 內。
Fig. 26.139 Field cells in acral melanoma: the top panel shows an acral melanoma with close-ups of four particular areas underneath. Area A shows superficially invasive melanoma that by FISH shows amplification of CCND1 (cyclin D1) and TERT telomerase reverse transcriptase (clusters of red and green signals in blue stained nuclei, respectively). Area B shows melanoma in situ with amplification of CCND1 and normal copy number of hTERT. Areas C and D show histopathologically uninvolved epidermis which by FISH harbors single basal melanocytes with amplification of CCND1 (clusters of red signals within blue stained nuclei). In the second FISH panel for area D a melanocyte with CCND1 amplification is situated in an eccrine duct.
它們反映了疾病的早期進展階段。一旦其臨床意義被界定,場效應 (field effect) 的判定相較於目前以經驗為基礎所推導的建議,或可為原發性黑色素瘤切除的手術邊緣 (surgical margins) 大小提供更客觀的指引。
雖然肢端與黏膜黑色素瘤同樣具有高發生率的染色體異常 (chromosomal aberrations),包括頻繁的基因擴增,但這些套數變異 (copy number alterations) 所影響的基因組區域,在兩者之間有顯著差異。肢端黑色素瘤中最常擴增的位點是位於 chromosome 11q13、編碼 cyclin D1 的 CCND1 基因座 (locus)。另一個常被擴增的位點包括位於 chromosome 5p15 的 TERT telomerase 基因座。然而,約有百分之五十的肢端黑色素瘤會擴增 CCND1,黏膜黑色素瘤則通常不會。取而代之的是,一部分黏膜黑色素瘤會擴增位於 chromosome 12q14、編碼 cyclin D1 結合夥伴的 cyclin dependent kinase 4 (CDK4)。CDK4 正常由 p16 的功能所抑制,p16 作為 G1/S 轉換點的守門員 (gatekeeper)。在黏膜黑色素瘤中,CDK4 的擴增與 p16 基因座的缺失 (deletions) 互斥 (mutually exclusive),顯示這些基因組異常在功能上可能相當。黏膜黑色素瘤亦具有 SF3B1 的反覆突變,此基因在葡萄膜黑色素瘤 (uveal melanoma) 中常見突變,而通常不見於肢端黑色素瘤。
尚待發現的機轉似乎對肢端與黏膜黑色素瘤的形成至關重要。
肢端與黏膜黑色素瘤中的此種染色體不穩定 (chromosomal instability) 發生於疾病進展的早期,甚至已在肢端原位黑色素瘤 (acral melanoma in situ) 周圍組織病理學上外觀正常的黑色素細胞中被偵測到,即所謂的場細胞 (field cells) 或場效應 (field effect)(Fig. 26.139)。場細胞帶有相鄰黑色素瘤中所見之部分(但非全部)基因擴增,顯示它們是組織病理學上明確之原位黑色素瘤的前驅 (precursors)(Fig. 26.140)。這些細胞代表了帶有嚴重基因變異(源自對基因組完整性維護之控制喪失)的黑色素細胞的純系擴增 (clonal expansions)。此類基因改變在性質上不同於 BRAF 等致癌基因 (oncogenes) 的點突變 (point mutations),因為它們意味著保護基因組完整性之保護因子的喪失。因此,場細胞最好被詮釋為原位黑色素瘤的早期型態或前驅。這些腫瘤性黑色素細胞的場 (fields) 可以相當廣泛,可出現於組織病理學可偵測之原位部分以外大於 1 公分之處,且場的大小與腫瘤厚度完全無關,更進一步顯示
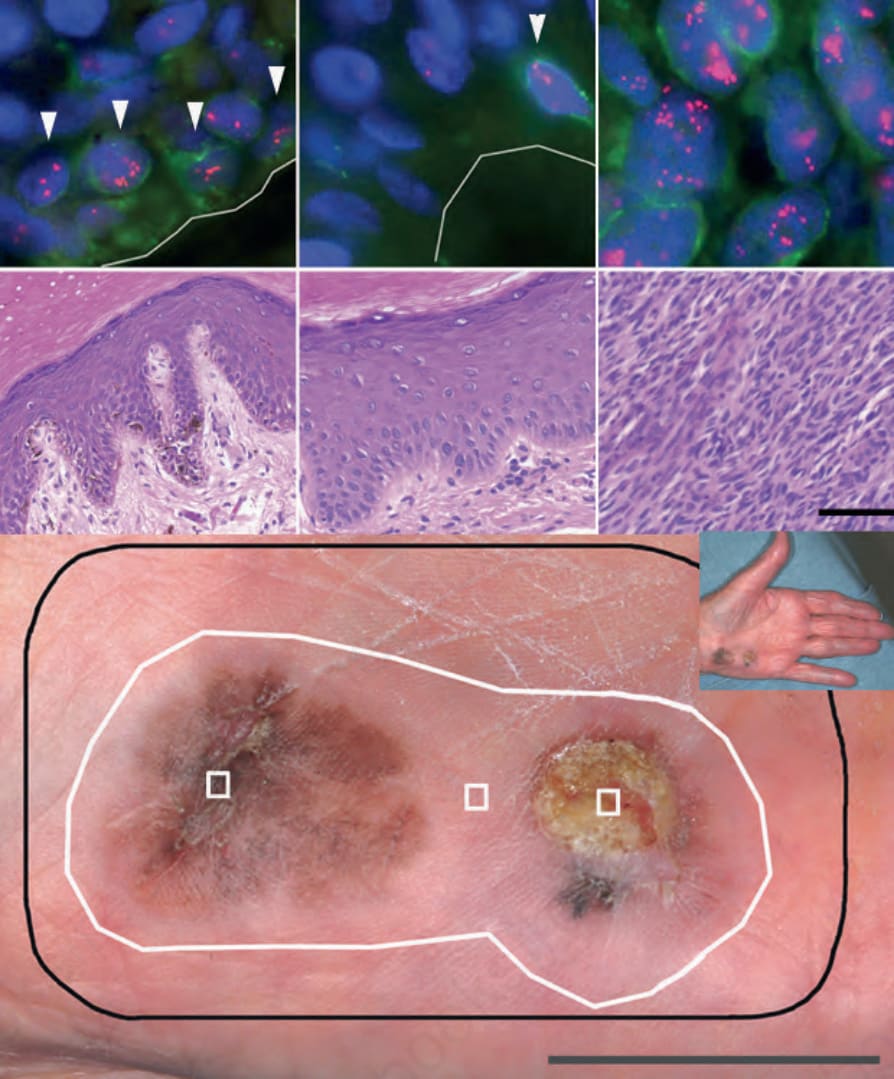
圖 26-140:肢端黑色素瘤中的場細胞 (field cells):下方面板:臨床圖像,包含 (1) 原位黑色素瘤 (melanoma in situ);(2) 臨床上、皮膚鏡上 (dermoscopically) 及顯微鏡下皆正常的皮膚;(3) 厚度 3.0 mm 的侵襲性黑色素瘤。黑線為手術邊緣 (surgical margin);白線為以 11q13 擴增所辨識的場細胞邊緣 (field cell margin)。中間與上方面板分別為區域 1–3 的 H&E 與 FISH。FISH 影像代表單一焦平面 (focal plane),故並非所有訊號皆可見。使用綠色免疫螢光標記的 Melan-A 抗體以協助辨識基底黑色素細胞。具 11q13 擴增的細胞於區域 1 與 2 以箭頭 (arrowheads) 標示(上方面板)。
Fig. 26.140 Field cells in acral melanoma: bottom panel: clinical picture with (1) melanoma in situ; (2) clinically, dermoscopically, and microscopically normal skin; (3) invasive melanoma of 3.0-mm thickness. Black line, surgical margin; white line, field cell margin identified by amplification of 11q13. Middle and upper panels, respectively: H&E and FISH for areas 1–3. FISH images represent one focal plane, so not all signals are visible. A green immunofluorescent labeled Melan-A antibody was used to aid in identifying basal melanocytes. Cells with amplification of 11q13 are highlighted by arrowheads for areas 1 and 2 (upper panels).
相較於肢端與黏膜黑色素瘤中大量的染色體套數變化與擴增,其突變負荷遠低於日光暴露黑色素瘤。許多突變呈現一種隨老化所見的突變生成 (mutagenesis) 模式,即 CpG 雙核苷酸 (dinucleotides) 處 5-methylcytosine 的自發性去胺基 (spontaneous deamination)。UV 標記突變大多不存在。肢端與黏膜黑色素瘤的多數遺傳研究聚焦於侵襲性黑色素瘤,而關於原位 (in situ) 階段的發表極少。肢端原位黑色素瘤中的 BRAFV600E 突變已有報導,但肢端黑色素瘤中 BRAF 突變的發生率(約百分之十五至二十)顯著低於低 CSD 黑色素瘤 (low-CSD melanomas)(約百分之七十),於黏膜黑色素瘤更低(約百分之五),顯示 BRAF 突變僅啟動了這些黑色素瘤中相對小的百分比。NRAS 突變以及 KIT 突變或擴增,較常見於肢端與黏膜黑色素瘤。這兩種突變會活化 MAP kinase pathway 並驅動增殖,因此合理地假設它們發生於肢端與黏膜部位之原位黑色素瘤的早期增殖期。未來研究有必要證實此一推測。TERT promoter 突變在肢端黑色素瘤中較不常見(約百分之十),但 TERT 基因擴增則頻繁可見,並已於肢端原位黑色素瘤中被記錄。
朝向未受侵犯的表皮方向。這些生長特性可能與 KIT 活化的生物學效應有關。KIT 對於黑色素母細胞 (melanoblast) 自神經嵴 (neural crest) 遷移至皮膚,以及在發育過程中歸巢 (homing) 至上皮結構,皆屬必要。若將組成性活化 (constitutively active) 的 KIT 在體外 (in vitro) 導入人類黑色素細胞,會誘發類似雀斑樣 (lentiginous) 模式的遷移表型 (migratory phenotype)。因此,黑色素瘤中異常的 KIT 訊息傳遞,或可協助解釋肢端皮膚黑色素瘤所描述的場效應(Figs 26.139 與 26.140)。
與上皮結構無關聯而發生的黑色素細胞腫瘤 (Melanocytic tumors arising without associations to epithelial structures)
大多數黑色素細胞腫瘤,包括良性黑色素細胞痣 (benign melanocytic nevi) 以及黑色素瘤,皆源自位於全身上皮結構內的黑色素細胞,並帶有如 BRAF、NRAS、NF1 與 KIT 等基因的突變。然而,一部分黑色素細胞腫瘤的發生與上皮結構無明顯連結,且不顯示這些突變。其中一類為葡萄膜黑色素瘤 (uveal melanoma),源自眼部脈絡叢 (choroidal plexus)、睫狀體 (ciliary body) 與虹膜 (iris) 內的黑色素細胞,在生物學上與皮膚黑色素瘤截然不同,具有極強的肝轉移傾向。它在某些染色體異常的存在上也有差異,例如頻繁的 chromosome 3 缺失,此於葡萄膜黑色素瘤中作為一個負向預後指標 (negative prognostic indicator)。在皮膚中,真皮內黑色素細胞增生 (intradermal melanocytic proliferations) 可為先天性或後天性,呈現多樣樣態,自離散的藍色丘疹(藍痣 blue nevi)到影響結膜與眼周皮膚的大片藍灰色斑(太田母斑 nevus of Ota)、肩部(伊藤母斑 nevus of Ito)以及下背部(蒙古斑 Mongolian spot)。真皮內黑色素細胞腫瘤與葡萄膜黑色素瘤之間潛在的關聯,由以下事實所提示:在白種人 (Caucasians) 中 nevus of Ota 是葡萄膜黑色素瘤的危險因子。此外,blue nevi 在細胞學上可顯示與葡萄膜黑色素瘤的相似性。有趣的是,雖然葡萄膜黑色素瘤在免疫組化 (immunohistochemistry) 上強烈表現 KIT,它們卻缺乏 KIT 基因的突變。
於小鼠的正向遺傳篩選 (forward genetic screen) 辨識出異三聚體 G 蛋白次單元 (heterotrimeric G-protein subunits) GNAQ 與 GNA11 的胚系突變,其藉由誘發黑色素細胞細微的真皮內增生而導致皮膚色素過度沉著 (hyperpigmentation)。GNAQ 與 GNA11 是 G 蛋白 α 次單元 q class 的成員,參與介導 G 蛋白偶聯受體 (G-protein coupled receptors, GPCRs) 與下游效應子 (downstream effectors) 之間的訊息傳遞。GNAQ 的體細胞突變 (somatic mutations) 見於百分之八十三的 blue nevi、百分之五十的類藍痣黑色素瘤(‘惡性藍痣’ ‘malignant blue nevi’)以及百分之四十六的葡萄膜黑色素瘤。人類腫瘤中的突變與小鼠篩選中所發現的基因變異不同。在人類黑色素細胞腫瘤中,所有突變皆專一地發生於 codon 209,此與 NRAS 家族成員的 codon 61 同源,並導致組成性活化。GNAQ 經由 phospholipase Cβ 釋放二醯甘油 (diacylglycerol, DAG) 而活化 protein kinase C (PKC) 家族成員 PKCδ 與 ε。體外研究顯示 GNAQQ209L 突變以與 NRAS 突變相當的效率轉化黑色素細胞,並導致 MAP kinase pathway 的活化。此 MAP kinase 活化是特異性地經由 DAG 將 RasGRP3 招募至細胞膜,並由 PKC δ 與 ɛ 將其磷酸化與活化而發生。RasGRP3 接著活化 RAS 以啟動 MAP kinase pathway 訊息傳遞。當導入小鼠黑色素細胞時,突變型 GNAQ 誘發出重度色素沉著的腫瘤,其形態類似人類 blue nevi 與色素性上皮樣黑色素細胞瘤 (pigmented epithelioid melanocytomas) 的譜系,證實其在驅動這些腫瘤型態中的角色。
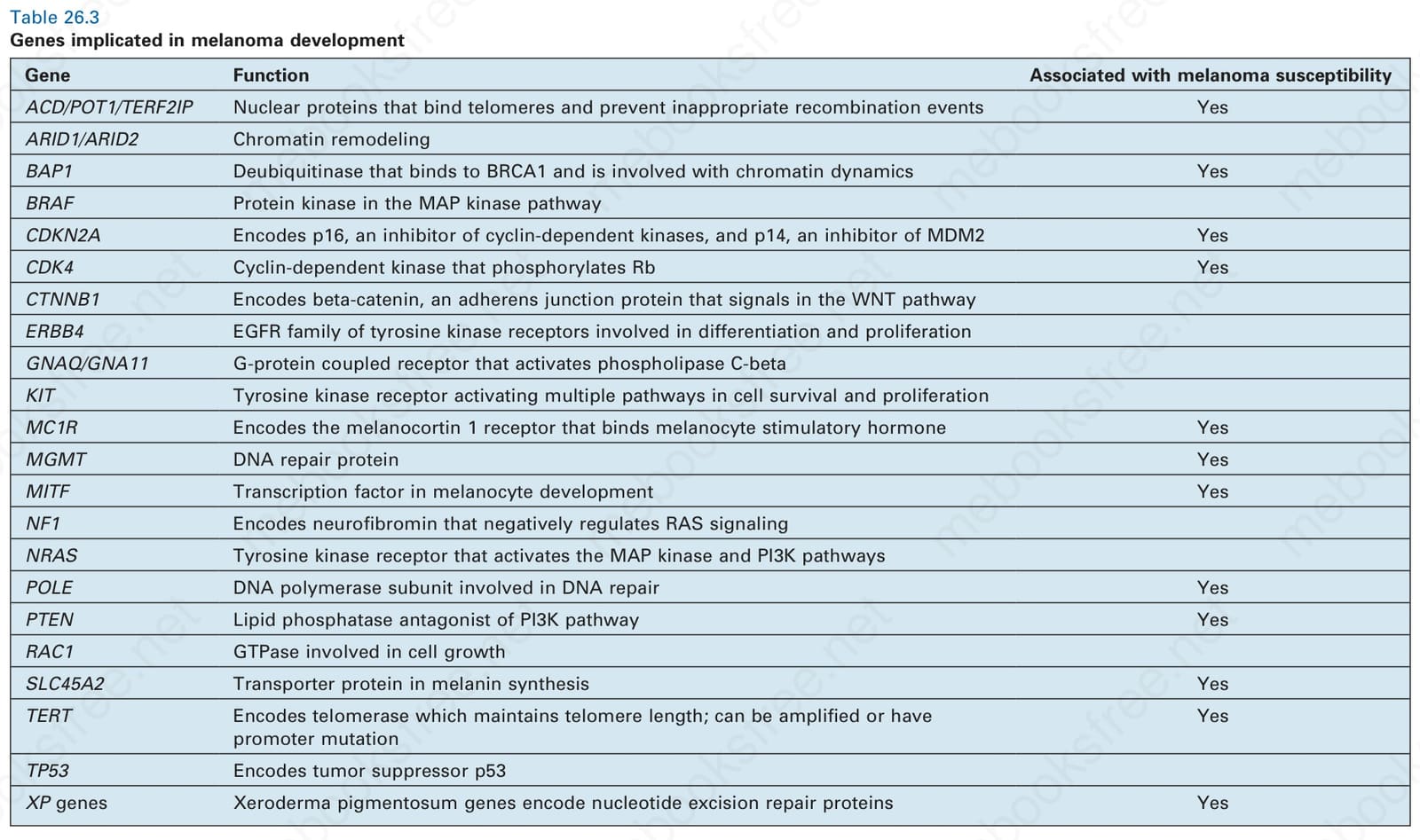
表 26-3:與黑色素瘤發展相關的基因 (genes implicated in melanoma development)。
Table 26.3 Genes implicated in melanoma development