Alopecia areata
Alopecia areata
Alopecia areata is the second most common form of alopecia in humans ranging from solitary foci of hair loss, loss of hair on the head, to total loss of all body hair. While the etiology has been debated for decades, it is now clearly established that this is a cell-mediated autoimmune disease.1 The C3H/HeJ mouse was discovered in 1991, 25 years ago, to develop patchy alopecia that waxed and waned but only occurred in 20% to 25% of the colony as they aged. The alopecia was initially observed in mice around 5 months of age, but the frequency in the colony rose and stabilized at around 12 months of age.2 Because lesions came and either expanded or disappeared, very similar to the human disease, and only a percentage of the mice came down with alopecia areata, the C3H/HeJ model proved difficult to work with as a preclinical or basic research tool. Discovery that full thickness skin grafts onto immunodeficient mice of the same strain resulted in normal hair regrowth, but grafts onto histocompatible mice resulted not only in retention of the alopecia phenotype but also development of progressive alopecia beginning in the axillary and inguinal regions, spreading across the abdomen and eventually to the back of the mice resulting in generalized alopecia. This yielded a highly reproducible mouse model that is heavily used today.3 Modifications of this protocol, most recently using draining lymph node derived cells, continue to be made.4
As with human alopecia areata, the late anagen and early catagen follicles are surrounded and infiltrated by a mixed inflammatory cell infiltrate, primarily consisting of CD4+ and CD8+ T cells. As with humans, CD8+ KLRK1 (killer cell lectin-like receptor C1 gene, previously called NKG2D) are the primary effector cells. The result is that there is a follicular dystrophy starting in the bulb region that leads to a deformity and weakness in
1910 Animal models of skin disease
remains in use today for xenografts, isografts, and allografts, especially in dermatology because the adults are alopecic.5 The alopecia evident in nude mice is due to a defect in the quality of their hair resulting in breakage as the hair shafts emerge from the hair follicles, and not that they do not have hair follicles.6 One has to be careful not to confuse nude mice with hairless mice (Hr), which also have no hair in adulthood but have a largely intact immune system such that they will not accept xenografts or allografts.
Many more mutant mice have been discovered or created which serve as improved hosts for human tissues, especially human cancers.7–9 Mice used for these types of studies are commonly called patient-derived xenograft (PDX) mice. Their value is that the cancers can be grafted from the patient to several recipient mice, allowed to grow, and then serially transplanted, allowing detailed molecular studies or testing with a variety of chemotherapeutic agents. The basic protocol involves mincing up fresh cancer biopsies into 3- to 5-mm3 pieces that are then surgically implanted subcutaneously into the lateral side of a NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ immunodeficient
mouse, commonly referred to as an NSG mouse. Mice are monitored for 5 months. Single cell suspensions or pieces of solid cancers that develop in these recipient mice can then be implanted into additional NSG mice to expand the numbers of mice carrying the cancers for research.10–12 A number of skin cancer PDX models are available to investigators at the time of this writing including 13 melanomas, 4 squamous cell carcinomas, and 1 Merkel cell tumor. Details can be found on these cancers in the Mouse Tumor Biology Database (http://tumor.informatics.jax.org/; 15 November 2016). A comparison between a human melanoma and the melanoma engrafted onto an NSG mouse is shown in Fig. 36.19; the histologic differences are negligible. Better immunodeficient mouse models are constantly being developed for these types of studies.13 Another useful mouse tool is to use mice that have a ubiquitously expressing marker, such as enhanced green fluorescent protein (EGFP), as the donor. Using either immunofluorescence or immunohistochemistry with antibodies specific for EGFP, it is possible to follow the lineages of the cells grafted, whether they came from the host or the donor.5
1911 Zebrafish as a model system to study skin biology and pathology
IMPC) programs, but even these lack critical expertise or depth in evaluation of all organ systems.
Two examples of diseases for which models are often thought not to exist include cutaneous scarring lesions/keloids and vascular tumors.
There are numerous models for cicatricial alopecia in mice1–3 in which scarring is limited to the hair follicles and are clearly distinct from keloids. However, a number of models are described in the literature that can be used for keloids, with most of them using PDX-type approaches.4–11
Vascular tumors are another disease group for which many believe mouse models do not exist. Cutaneous vascular tumors do arise spontaneously in mice, albeit very rarely. However, they are relatively common in many inbred strains but only in very old mice.12–15 When one performs a literature search (by including terms such as hemangioma, hemangiosarcoma, and hemangioendothelioma), not surprisingly, a number of models emerge, again often based on the PDX approach.16–24
There are huge numbers of mouse models for most diseases already available. More are under development. As investigators continue to refine existing models, molecular pathways, or with genetic modifications, addition of more than one mutation, changing strain background, or by simply utilizing more refined and systematic phenotyping methods, better models will continue to emerge. Large biorepositories, such as the one at the Jackson Laboratory (JAX), make these mouse models available to investigators worldwide.
A C E
B D F
Zebrafish as a model system to study skin biology and pathology
Several animal models have been developed to recapitulate the features of specific skin diseases, and such model systems have provided valuable insight into the pathomechanistic details of these diseases. They have also provided useful systems for testing of treatment modalities for a number of conditions. The preferred platform for such model development has been the mouse, often through the development of ‘knockout’ mice or by development of transgenic mice with expression of the mutant genes. Although the mouse models have demonstrated remarkable similarity to human diseases, mice as a model system have considerable limitations, including relatively long life span, extended reproductive cycle, and high cost of development and housing. Also, the genetic background of the mouse strains can have a major influence on the development of the disease phenotype. In some cases, development of the mouse model is not feasible because of the absence of the corresponding gene in the mouse genome. These considerations, particularly in conjunction with cost-containment issues, have prompted the search for an alternative model system to study skin diseases. In fact, Vanchieri1 suggested, ‘Move over mouse: make way for the woodchucks, ferrets, and zebrafish’, and Lieschke and Currie2 said, ‘Animal models of human disease: zebrafish swim into view’.
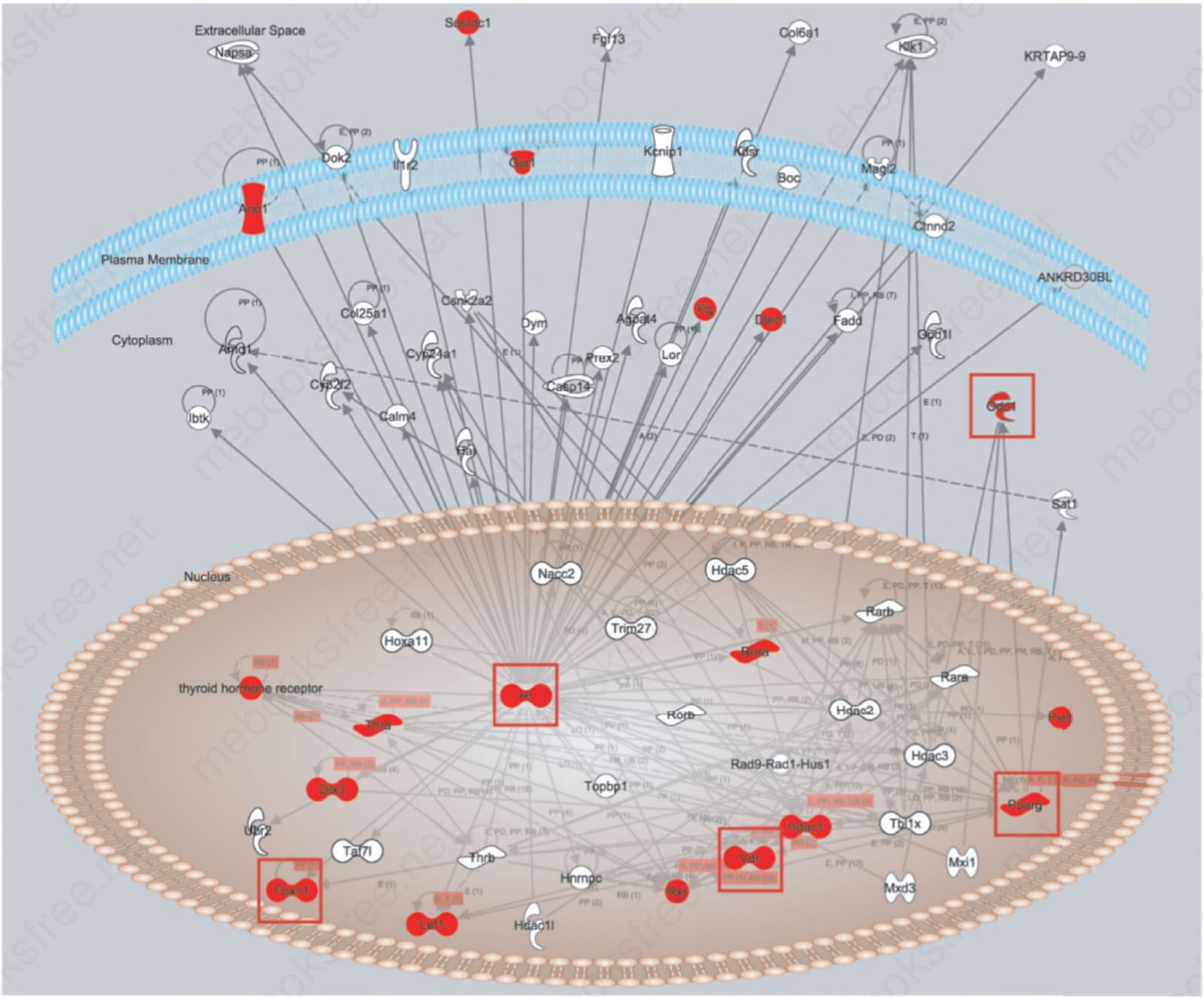
Fig. 36.16 Ingenuity Pathway Analysis Software® (http://www.ingenuity.com/) molecular pathway diagram for genes that interact with hairless (HR). Mutations in both mice and humans in the Hr/HR and Vdr/VDR genes result in genocopies, the same phenotypes in both groups of mutants. As these interact with other genes, it is not surprising that follicular dystrophy results, although the phenotype is different, as with FOXN1 (nude human or mouse) or DLX3.
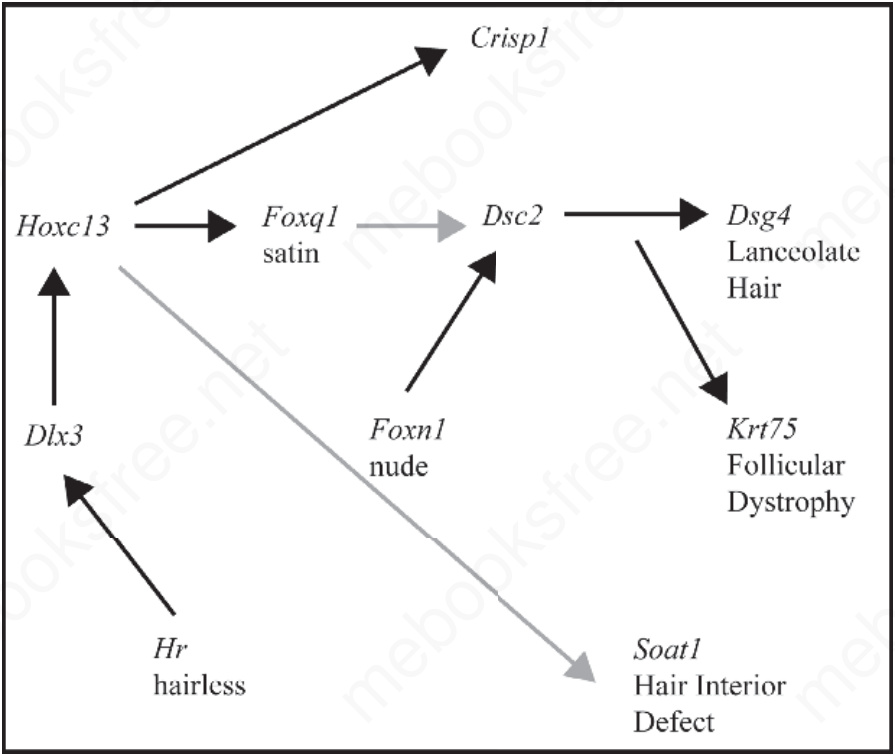
Fig. 36.17 Known and predicted molecular pathway for hair follicle development. Mice with mutations in these genes develop skin lesions that include various forms of follicular dystrophy. Some, such as Soat1 and Foxq1, result in defects within hair shafts, whereas others, such as Hoxc13 and Foxn1, result in weak, distorted shafts that break off. Integrating these individual mouse models for each gene in the pathway enables researchers to define their interactions and follow how defects cause clinical disease.
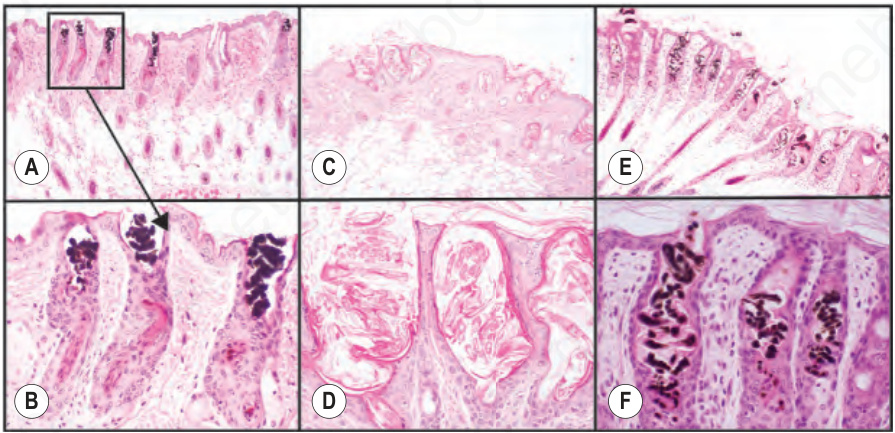
Fig. 36.18 C57BL/6J-Hoxc13tm1Mrc null adult mice (A, B) have weak hair shafts that twist within the infundibulum and break off at the skin surface. Nude mice (C, D; CByJ. Cg-Foxn1nu albino; E, F; B6.Cg-Foxn1nu, pigmented) have a similar phenotype. These genes work in the same molecular pathway.
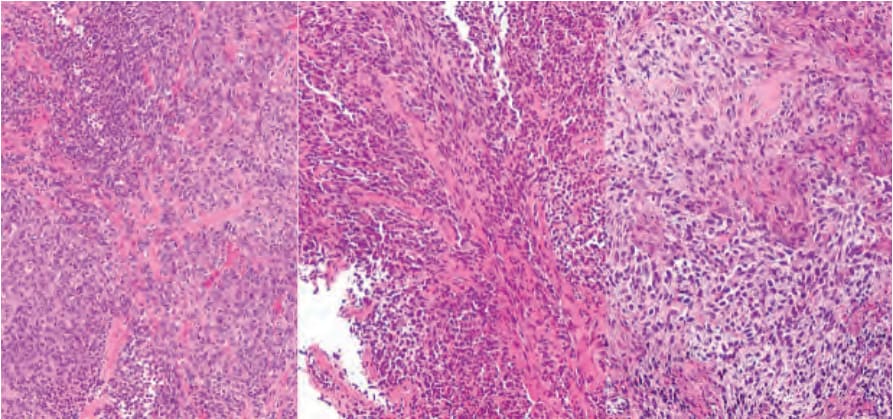
Fig. 36.19 Patient-derived xenograft (PDX) melanoma model. The left image shows the histology of a melanoma in a human patient while the two images on the right show the same neoplasm as two independent PDX in two separate immunodeficient mice. Images courtesy of Clemens Krepler and James Hayden, The Wistar Institute, Philadelphia, PA and Xiaowei (George) Xu, University of Pennsylvania, Philadelphia, PA.