Intermediate tumors (dysplastic/atypical nevi)
Intermediate tumors (dysplastic/atypical nevi)
The existence of an intermediate category between the spectrum of nevus and melanoma has been controversial. Diagnostic discordance in tumors with mixed histopathological features has been documented in numerous studies, indicative of a histopathological gray zone but not necessarily a biological one. A recent investigation into the genetic events associated with melanoma progression has characterized the genetic profile of such intermediate lesions with intermediate histopathological characteristics.20 Unequivocal nevi, defined in the study by diagnostic concordance among multiple dermatopathologists, were found to have a single driver mutation, typically BRAFV600E. By contrast, intermediate lesions with more atypical histopathological features also had activating mutations in the MAP-kinase pathway, but these were enriched for other types of BRAF mutations and NRAS mutations. Intermediate lesions typically also had additional mutations such as TERT promoter and/or hemizygous CDKN2A mutations, supporting the existence of a genetically distinct intermediate category in which multiple genetic aberrations have accrued rather than the solitary pathogenic mutations found in unequivocal benign nevi. The transition to invasive melanoma is characterized by further genetic aberrations and includes mutations in genes such as PTEN, TP53, ARID1/2, and homozygous CDKN2A mutations. Indications of intermediate stages have also been found in neoplasms with a cellular blue nevus phenotype38 and atypical Spitz tumors.39,40
1309 The molecular pathology of melanocytic nevi
lesion as malignant just because one barrier is dysfunctional. Assessment of cell proliferation also does not determine whether such proliferative activity within a given lesion is temporary, i.e., pre-senescent, or unrestricted.
The ability to detect clonal populations of cells with chromosomal aberrations represents a diagnostic parameter to assess the aftermath of barrier failure. CGH studies of benign nevi of various types show that the vast majority have no detectable copy number changes.41 This finding does not exclude the possibility that chromosomal aberrations exist in individual melanocytes of nevi or that random aberrations are present in many of the cells. This finding indicates that any such cells have not undergone significant clonal expansion. Only a few categories of benign nevi studied by CGH have registered copy number changes. These include gains of chromosome 11p in HRAS mutant Spitz nevi44 and gains or losses of entire chromosomes in atypical nodular proliferations (proliferative nodules) arising in giant congenital nevi.45 Copy number increases of chromosome 11p are not found in melanoma, and the aberrations in atypical nodular proliferation also have distinctive features. The latter frequently show gains or losses of entire chromosomes including loss of chromosome 7, which if altered in melanoma is gained, and chromosome 10 gain, which shows reciprocal copy number changes in melanoma. In contrast to the whole chromosomal gains and losses of proliferative nodules, most melanomas have losses or gains involving partial chromosomes. The most common aberrations in melanoma in decreasing order of frequency include losses of chromosomes 9p, 10q, 9q, 6q, 8p, and 11q and gains of chromosome 6p, 7, 1q, 8q, 17q, and 20 (Fig. 25.262). Therefore, both detection of aberrations and the pattern of aberrations help distinguish between nevi and melanomas.
Striking differences exist in the number of genetic aberrations such as chromosomal gains and losses between nevi and melanomas. While the vast majority of melanomas show chromosomal aberrations, they are infrequent in nevi (see Fig. 25.262).41 These chromosomal aberrations in melanomas are clonal in nature, i.e., aberrations that are found in the majority of the constituent melanocytes.42 This can be demonstrated by techniques such as CGH, which measures the average DNA copy number of a population of cells across the entire genome.43 Additionally, the pattern of chromosomal gains and losses is not random, but preferentially selects for specific chromosomal regions that harbor critical genes that inhibit progression (and therefore are decreased in copy number) or promote melanoma growth (and therefore are increased in copy number). For example, chromosome 9p21, which harbors the CDKN2A gene mentioned above, is lost in more than 60% of melanomas, whereas chromosome 7q31, where BRAF is located, is commonly gained, in particular in melanomas with BRAF mutations.
The recurrent pattern of chromosomal aberrations in melanoma allows for assessment of such aberrations by more focused methods such as FISH. FISH can be applied to fixed tissue and allows a faster turnaround time and analysis of smaller samples than CGH, including entirely intraepidermal lesions. Multiple studies have shown the combination of probes targeted to chromosomes 6p, 6 centromere, 6q, and 11q can distinguish melanomas from nevi with a high specificity (approximately 95%) and acceptable sensitivity (approximately 85%).46–49 The sensitivity and specificity in histopathologically ambiguous tumors is more difficult to assess given the limited number of such cases with long-term follow-up data. Of particular interest are spitzoid tumors, which are particularly prone to diagnostic uncertainty. The limited available data on such tumors that eventuated in metastatic disease and death suggest that homozygous CDKN2A loss and TERT promoter mutation are associated with metastatic potential.40,50 A second set of FISH probes targeting chromosomes 6p, 8q, 9p, and 11q has been adopted by some to better target spitzoid tumors.51 The use of tailored probe sets for specific differential diagnostic situations could help raise the sensitivity of FISH-based assays in the future.
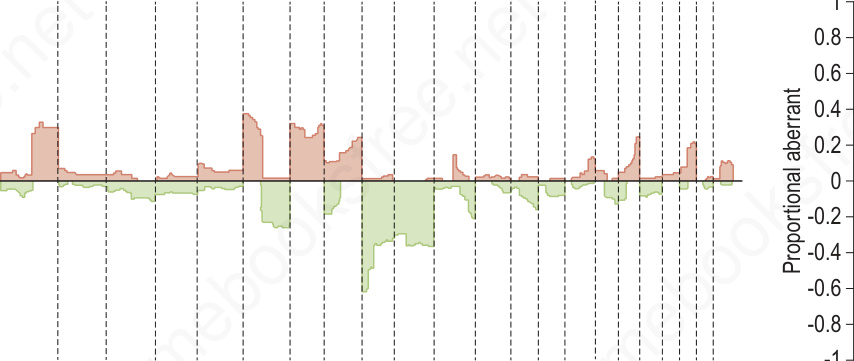
Fig. 25.262 Comparative genomic hybridization of melanoma (malignant) and nevi (benign): on the left, melanomas (n = 133) are associated with multiple copy number gains and losses that cluster. In contrast, the nevi (n = 54) on the right show minimal changes with the exception of the 11p copy number increase or amplification in Spitz nevi which includes the HRAS gene. These differences can be exploited using multiplexed FISH assays to support the diagnosis of melanoma or nevus in challenging cases.