黑色素瘤的分子分類 — 第二部分 (Molecular Classification of Melanoma — Part 2)
痣演進為黑色素瘤的分子背景
- 上述後者的突變反映出複製性老化 (replicative senescence) 與維持 G1/S 檢查點 (G1/S checkpoint) 控制,在防止痣 (nevi) 演進為黑色素瘤 (melanoma) 上的重要性。UV signature mutation 圖譜將紫外線輻射 (UV radiation) 標示為痣轉化為黑色素瘤之過程中的驅動因子。
低累積日照損傷之黑色素瘤 (Melanoma with low cumulative sun damage, low-CSD)
- low-CSD 黑色素瘤最常發生於 50 歲以下的患者,位於軀幹或四肢,組織病理上呈現低至中度的日光彈性纖維變性 (solar elastosis)。
- BRAFV600E 突變在 low-CSD 黑色素瘤中極為常見(約 70%),為主要的驅動突變 (driver mutation),其次為 NRAS 突變(約 20%)。
- BRAF 突變型黑色素瘤常表現出特定的組織形態學特徵,例如表皮內黑色素細胞向上散布增加(pagetoid scatter,佩吉特樣散布)、巢狀結構 (nests) 多於單一細胞、受累表皮增厚、與鄰近未受累表皮間有更銳利的分界,以及構成腫瘤的細胞較大、較圓、且色素更深(Fig. 26.134)。
- 在一項研究中,將這些彼此獨立相關的特徵作簡單組合,可以 90% 的準確度預測 BRAF 突變狀態。這些組織形態學標準在預測突變狀態上的表現也優於傳統的 WHO 黑色素瘤分類,顯示出發展更佳分類方案的潛力——可定義出生物學上更為均質的亞群。隨著針對特定基因改變(包括 BRAF inhibitors 與 KIT inhibitors)之標靶治療的使用日益增加,此議題更具關聯性。
由原位到侵襲性黑色素瘤的演進
- 由原位 (in situ) 到侵襲性 (invasive) 黑色素瘤的演進,涉及在表皮範圍之外不受限制生長所需的額外基因改變。
- 單憑能在真皮內生長與存活並不代表已發生惡性轉化,因為黑色素細胞痣 (melanocytic nevi) 的細胞本已具備此特性。
- 細胞週期檢查點 (cell cycle checkpoint) 調控的喪失,對於釋放或增加侵襲性黑色素瘤相關的增殖能力至關重要,此點可由原位轉變為侵襲性黑色素瘤過程中,編碼 p16 與 p14ARF 的 CDKN2A 基因頻繁發生同型合子缺失 (homozygous loss) 得到佐證。
- 編碼關鍵腫瘤抑制蛋白(如 TP53 與 PTEN)之基因的額外突變,則發生於演進至更晚期疾病的較後階段。此外,侵襲性黑色素瘤必須透過誘導免疫耐受 (immune tolerance) 等機制,逃避抗腫瘤免疫監視 (antitumor immune surveillance)。
與前驅痣的關聯
- 黑色素細胞痣與 low-CSD 黑色素瘤共有的高比例 BRAFV600E 突變,支持兩者在紫外線輻射與曝曬時機上具有共同病因。
- 約四分之一至三分之一的黑色素瘤起源於組織病理上可辨識的前驅痣 (precursor nevi),而 low-CSD 黑色素瘤是其中最主要可辨識出前驅痣的黑色素瘤類型。
- 通常與相關痣並存的僅有黑色素瘤原位 (melanoma in situ),而後者(痣)在底下的真皮中仍可辨識。melanoma in situ 帶有與前驅痣相同的起始突變(通常為 BRAF 或 NRAS),並伴隨額外的基因異常,如 TERT promoter 突變或異型合子 (heterozygous) CDKN2A。
基因組不穩定性與端粒
- 絕大多數黑色素瘤都表現出某種程度的基因組不穩定性 (genomic instability),形式為染色體的增益 (gains) 與缺失 (losses)。此不穩定性似乎與良性黑色素細胞痣轉變為 melanoma in situ 的過程同時發生,因為痣傾向於沒有染色體拷貝數變化,而 melanoma in situ 中則可偵測到。
- 此不穩定性可能與端粒縮短 (telomere shortening),以及端粒受侵蝕的姊妹染色分體 (sister chromatids) 發生端對端融合 (end-to-end fusion)、產生在有絲分裂 (mitosis) 中可能斷裂的雙著絲點染色體 (dicentric chromosomes) 有關。
- TERT promoter 突變在黑色素瘤演進早期所具有的選擇性優勢,顯示複製性老化在轉化過程中成為一道早期屏障。TERT promoter 突變僅導致端粒酶 (telomerase) 表現的些微提升,這雖能保住臨界縮短的端粒,卻無法完全抑制端粒融合,因而產生不穩定性、generate DNA copy number 變化。這也解釋了為何帶有 TERT promoter 突變之黑色素瘤的端粒仍維持短小。
各亞型間染色體增益與缺失的差異
- 雖然各黑色素瘤亞型間染色體增益與缺失的模式有些相似之處,但依紫外線曝曬程度區分的腫瘤之間,仍存在顯著差異。
- 染色體 6p 的增益與 6q 的缺失在 low-CSD 與 high-CSD 黑色素瘤中皆常見,但染色體 10 的缺失發生於 40% 的 low-CSD 黑色素瘤、卻不到 10% 的 CSD 黑色素瘤(Fig. 26.135)。
- 染色體 10 上常見缺失的區域含有腫瘤抑制基因 PTEN,其編碼 PI3 kinase 路徑的負向調節因子,被認為是染色體 10 缺失的主要選擇壓力。BRAF 突變僅活化 MAP kinase 路徑,這解釋了為何同時併有 PTEN 缺失(以活化 PI3 kinase 路徑)具有優勢(Fig. 26.136)。
- 相對地,NRAS 的活化突變同時活化 MAP kinase 與 PI3 kinase 路徑,因此在帶有 NRAS 突變的腫瘤中無需 PTEN 缺失。PI3 kinase 催化次單元 (catalytic subunit) 的點突變 (point mutations) 也曾在少數黑色素瘤中被發現。
增殖與存活路徑的活化
- 染色體 7 的增益在 low-CSD 黑色素瘤中較為常見。BRAF 位於 7q34,其拷貝數增加反映了突變型 BRAF 等位基因 (allele) 的重複,提供增殖優勢。
- 位於染色體 11q13、編碼 cyclin D1 的 CCND1 拷貝數增加,是另一項區別特徵——其在 low-CSD 黑色素瘤中發生頻率低,卻存在於高達 50% 的 high-CSD 黑色素瘤中。
- 這些基因變化展示了在黑色素瘤演進過程中,主要的增殖與存活路徑如何同時被活化。這可透過不同的突變與染色體異常組合來達成,例如上游酪胺酸激酶受體(如 KIT 或 NRAS)的活化突變同時活化 MAP kinase 路徑與 PI3K 路徑,或透過 BRAF 的活化突變併合 PTEN 缺失(Fig. 26.136)。
高累積日照損傷之黑色素瘤 (Melanoma with high cumulative sun damage, high-CSD)
- high-CSD 黑色素瘤通常發生於 50 歲以上的白人 (Caucasians),多位於頭頸部,組織病理上呈現明顯的日光彈性纖維變性 (solar elastosis)。
- 與 low-CSD 黑色素瘤相對照,high-CSD 黑色素瘤通常沒有相關的前驅痣,而是在表皮內「de novo(從頭)」發生。
- 這類黑色素瘤常以延長的原位階段為特徵,腫瘤在轉為侵襲性之前,可歷經多年生長至數公分大小。關於 high-CSD 黑色素瘤原位階段的基因組資料有限。
- 約 20% 的此類原位黑色素瘤帶有 BRAF 突變,惟在此群中,BRAFnon-V600E 突變(例如 BRAFV600K)比 BRAFV600E 突變更為常見。
- 除了 high-CSD 與 low-CSD 黑色素瘤之間 BRAF 突變上的顯著差異(15% 對 70%)之外,也發現了其他基因差異。例如,有些研究報告高達 30% 的 high-CSD 黑色素瘤帶有 KIT 的突變或 DNA 拷貝數增加,而 KIT 突變在 low-CSD 黑色素瘤中罕見(Fig. 26.137)。NRAS 突變頻率的差異曾有部分研究提出,但似乎並非一致的發現。
促結締組織增生性黑色素瘤 (Desmoplastic melanoma)
- desmoplastic melanoma 是發生於慢性日光損傷皮膚的一種特定變異型黑色素瘤,具有高突變負荷 (mutation burden),通常超過所有其他黑色素瘤(中位數為 62 mutations/Mb,相較於 low-CSD 黑色素瘤約 15/Mb)。
- desmoplastic melanoma 的突變圖譜具有強烈的紫外線輻射特徵,其顯著之處在於完全缺乏 BRAF 與 NRAS 熱點突變 (hotspot mutations),並有高比例的去活化型 (inactivating) NF1 突變,後者在其上覆的原位成分中亦可偵測到。
- 此外,還可見 NFKBIE promoter 的反覆突變(NFKBIE 編碼一種核因子 NF-κB 信號的抑制因子),以及 TP53、CDKN2A、ARID1A、ARID2 與 RB1 的去活化型突變。TERT promoter 突變在 desmoplastic melanoma 中也很常見。
- desmoplastic melanoma 的染色體拷貝數改變較大多數其他黑色素瘤少,但仍可見癌基因(如 EGFR、CDK4、CCND1、MDM2、TERT 與 MAP3K1)的局部擴增 (focal amplifications),以及腫瘤抑制基因(如 CDKN2A)的局部缺失。
紫外線受保護部位(肢端與黏膜)之黑色素瘤
- 〔接續上文〕角質層 (corneum) 提供額外的紫外線保護。甲基質 (nail matrix) 受近端甲褶 (proximal nail fold) 與甲板 (nail plate) 保護。除了唇部與球結膜 (bulbar conjunctiva) 之外,黑色素瘤發生的黏膜部位並不暴露於日光。
- 對來自這些紫外線受保護部位之黑色素瘤進行 CGH 與 DNA 定序的基因組分析顯示,其呈現高度的基因組不穩定性,反映於頻繁的染色體異常,尤其是聚焦性的基因擴增 (focused gene amplifications),以及低突變負荷(Fig. 26.138)。
- Fig. 26.138 中的黏膜範例尤其顯示出影響幾乎每一條染色體的拷貝數變化。雖然黏膜黑色素瘤 (mucosal melanomas) 通常於相當大的延遲後才被偵測到,因而傾向比其他黑色素瘤類別的原發病灶更厚,但此高度的基因組不穩定性在較薄的病灶中即可顯示出來。
- 在其他癌症中,基因擴增通常出現於演進的晚期,且往往是不良預後的徵象。相對地,在肢端黑色素瘤 (acral melanoma) 中,這些擴增在演進的極早期即出現,於原位階段即可偵測到。
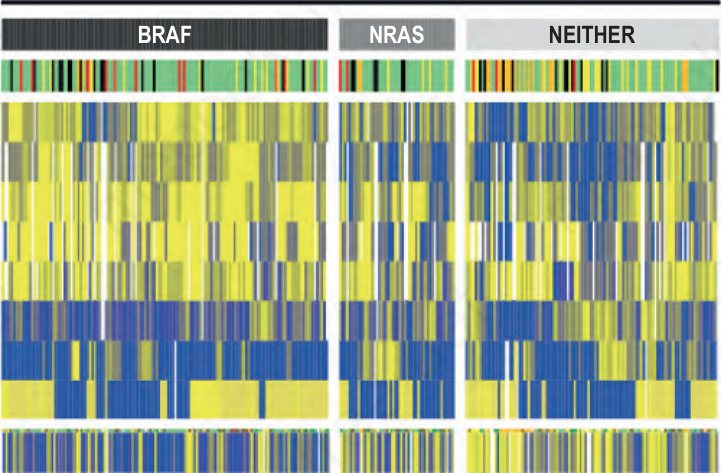
圖 26-134:依突變狀態呈現 302 例原發性黑色素瘤的形態學與臨床特徵:此熱圖 (heat map) 顯示與三組黑色素瘤——BRAF 突變型、NRAS 突變型或皆無突變——之突變狀態顯著相關的特徵。下列各列特徵的分數範圍由藍色色調(低分)經灰色(中間分數)至黃色(高分):腫瘤性黑色素細胞的色素沉著、表皮內黑色素細胞的向上散布、黑色素細胞的巢狀化、日光彈性纖維變性的程度、細胞大小與患者年齡。就境界 (circumscription) 而言,黃色代表由受累至鄰近未受累皮膚的突然轉變、灰色分數代表連續轉變、藍色分數代表不連續轉變。表皮輪廓 (epidermal contour) 由黃色(棘層肥厚 acanthotic)至藍色(萎縮 atrophic,指受黑色素瘤侵犯的表皮)。細胞形狀由藍色(圓形 round)至黃色(梭形 spindled)。樣本以欄列出,並於各類別內以聚合式階層聚類 (agglomerative hierarchical clustering) 排序。WHO 類型的色碼為:表淺擴散型黑色素瘤 (superficial spreading melanoma, SSM) 綠色、惡性雀斑樣痣黑色素瘤 (lentigo maligna melanoma, LMM) 黃色、結節型黑色素瘤 (nodular melanoma, NM) 紅色、肢端雀斑樣痣黑色素瘤 (acral lentiginous melanoma, ALM) 橘色。無法分類的樣本為黑色。 (Fig. 26.134)
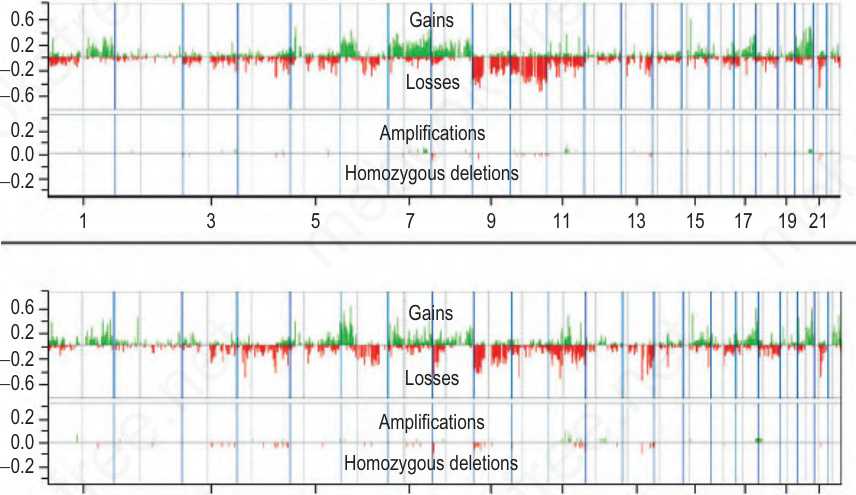
圖 26-135:non-CSD 與 CSD 黑色素瘤中 DNA 拷貝數變化的頻率:每種黑色素瘤類型的上方直方圖顯示增益 (gains,綠色) 與缺失 (losses,紅色) 的頻率,下方直方圖顯示擴增 (amplifications,綠色) 與同型合子缺失 (homozygous deletions,紅色)。x 軸代表由染色體 1 至 22 的基因組位置。垂直實心藍線標示染色體之間的邊界,垂直虛灰線標示著絲點 (centromeres) 的位置。 (Fig. 26.135)
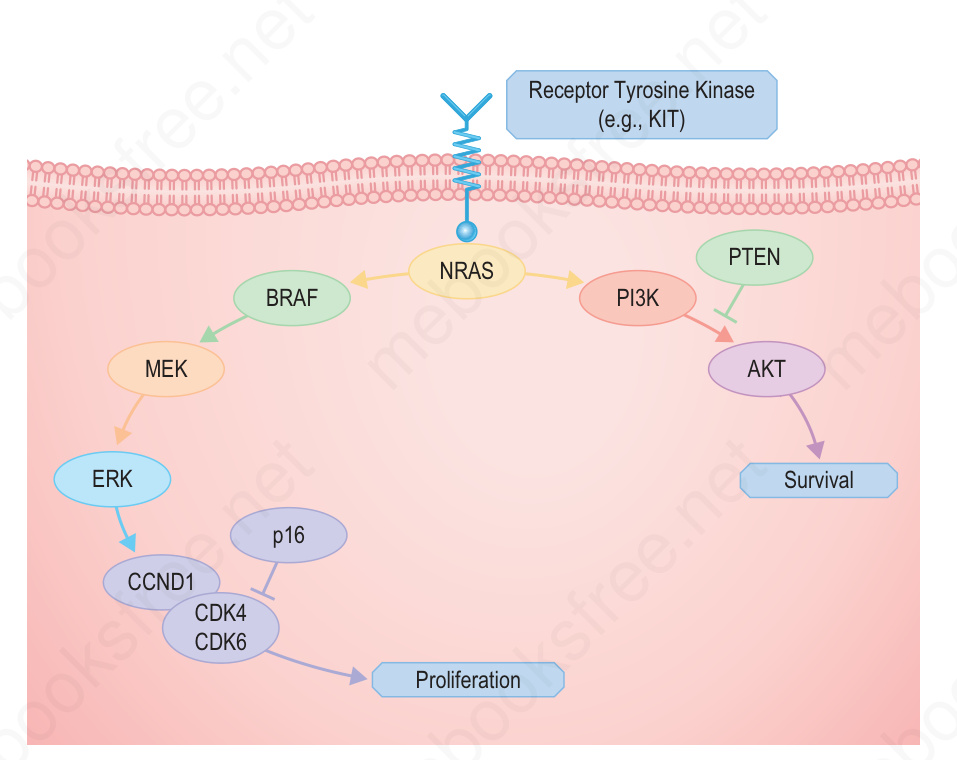
圖 26-136:黑色素瘤中重要路徑的簡化示意圖:某些受體酪胺酸激酶 (receptor tyrosine kinases) 的下游為 Ras,其分歧為 BRAF/ERK 與 PI3K/AKT 路徑,導向增殖 (proliferation) 與存活 (survival)。黑色素瘤的發生似乎需要這兩條(及其他)路徑皆被活化,並以合理方式加以分組。例如,KIT、NRAS 與 BRAF 的活化突變彼此互斥 (mutually exclusive),因為它們活化相同的路徑;而 PTEN 缺失見於 BRAF 突變、不見於 NRAS 突變,因為 NRAS 會活化 PI3K 路徑而 BRAF 不會。隨著更具特異性的抑制劑被開發出來,這些組合很可能具有治療上的意義。 (Fig. 26.136)

圖 26-137:四組黑色素瘤中 BRAF、NRAS 與 KIT 基因改變的頻率分布:non-CSD,無慢性日光誘發損傷之皮膚上的黑色素瘤;CSD,有慢性日光誘發損傷(以明顯日光彈性纖維變性為證)之皮膚上的黑色素瘤;acral,足底、手掌或甲下部位的黑色素瘤;mucosal,黏膜上的黑色素瘤。一例 CSD 黑色素瘤同時帶有 KIT 與 NRAS 突變,一例 acral 黑色素瘤同時帶有 KIT 與 BRAF 突變。 (Fig. 26.137)
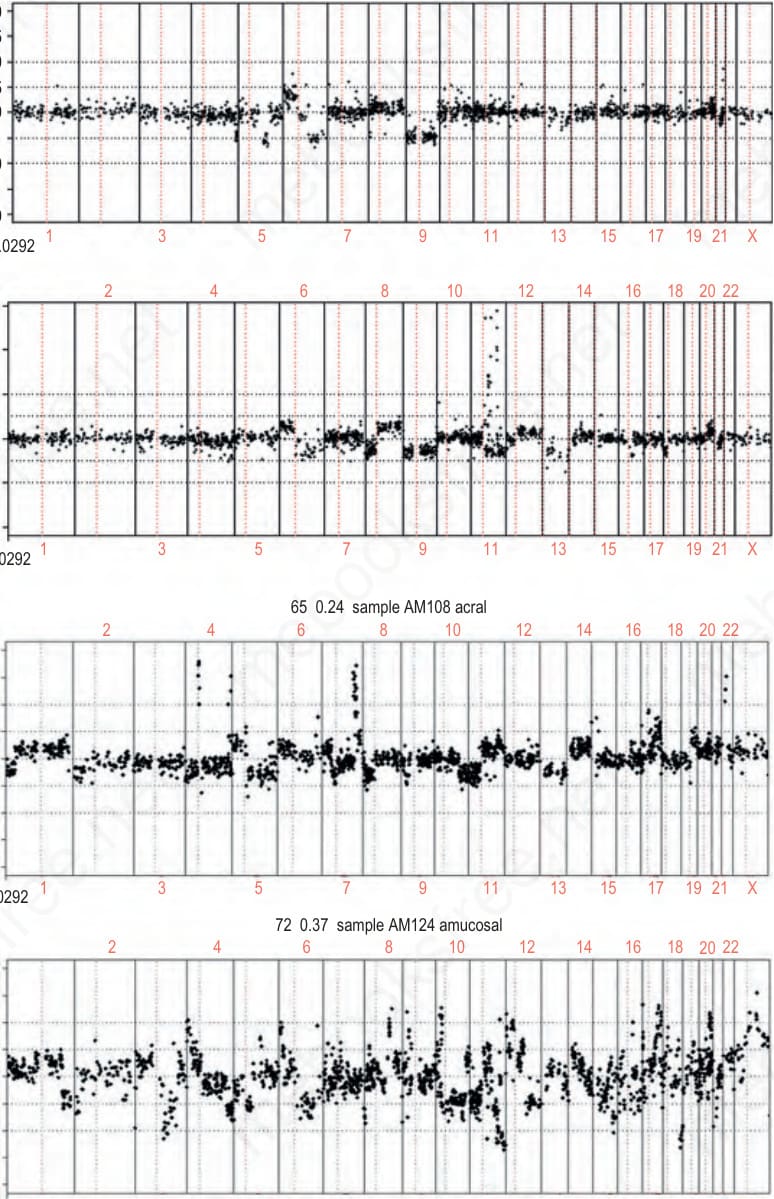
圖 26-138:黑色素瘤類型間基因組不穩定性程度的差異:non-CSD、CSD、acral 與 mucosal 黑色素瘤之 array CGH 圖譜的代表性範例。y 軸代表每個陣列元素 (array element) 的拷貝數(三重測量的平均值),以腫瘤對參照螢光強度比值的 log2 表示。介於 +0.25 與 –0.25 之間的數值視為正常;高於此範圍者為增益,低於此範圍者為缺失。高於 0.9 的數值視為擴增。實務上採用更複雜的評估,將每個個別病例的訊號雜訊比 (signal-to-noise ratio) 納入考量。x 軸代表由染色體 1p 至 22 的基因組位置。 (Fig. 26.138)
與黑色素瘤相關之基因功能與易感性
| 基因 | 功能 | 與黑色素瘤易感性相關 |
|---|---|---|
| ACD/POT1/TERF2IP | 結合端粒並防止不當重組事件的核蛋白 (Nuclear proteins that bind telomeres) | 是 |
| ARID1/ARID2 | 染色質重塑 (Chromatin remodeling) | |
| BAP1 | 結合 BRCA1、參與染色質動態的去泛素酶 (Deubiquitinase) | 是 |
| BRAF | MAP kinase 路徑中的蛋白激酶 (Protein kinase) | |
| CDKN2A | 編碼 p16(cyclin-dependent kinases 抑制因子)與 p14(MDM2 抑制因子) | 是 |
| CDK4 | 磷酸化 Rb 的 cyclin-dependent kinase | 是 |
| CTNNB1 | 編碼 beta-catenin,一種在 WNT 路徑中傳遞信號的黏附連接 (adherens junction) 蛋白 | |
| ERBB4 | 參與分化與增殖的 EGFR family 酪胺酸激酶受體 | |
| GNAQ/GNA11 | 活化 phospholipase C-beta 的 G 蛋白偶聯受體 (G-protein coupled receptor) | |
| KIT | 活化細胞存活與增殖中多條路徑的酪胺酸激酶受體 | |
| MC1R | 編碼結合黑色素細胞刺激激素 (melanocyte stimulatory hormone) 的 melanocortin 1 receptor | 是 |
| MGMT | DNA 修復蛋白 (DNA repair protein) | 是 |
| MITF | 黑色素細胞發育中的轉錄因子 (Transcription factor) | 是 |
| NF1 | 編碼負向調節 RAS 信號的 neurofibromin | |
| NRAS | 活化 MAP kinase 與 PI3K 路徑的酪胺酸激酶受體 | |
| POLE | 參與 DNA 修復的 DNA polymerase 次單元 | 是 |
| PTEN | PI3K 路徑的脂質磷酸酶 (lipid phosphatase) 拮抗劑 | 是 |
| RAC1 | 參與細胞生長的 GTPase | |
| SLC45A2 | 黑色素合成中的轉運蛋白 (Transporter protein) | 是 |
| TERT | 編碼維持端粒長度的 telomerase;可被擴增或帶有 promoter 突變 | |
| TP53 | 編碼腫瘤抑制因子 p53 | 是 |
| XP genes | Xeroderma pigmentosum 基因,編碼核苷酸切除修復 (nucleotide excision repair) 蛋白 | 是 |
表 26-x:與黑色素瘤易感性相關之基因及其功能 (genes associated with melanoma susceptibility)。