8.5hpf
8.5hpf
8.5 hpf 8.5 hpf B A
transmission electron microscopy revealed perturbations in the BMZ (Fig. 36.23A–B), mimicking features in patients with a form of junctional epidermolysis bullosa due to mutations in the human COL17A1 gene. The abca12 morphant fish demonstrated noticeable changes in the distribution of pigment along the trunk and tail. Scanning electron microscopy demonstrated the absence of microridges and development of scale-like spicules on the surface of the skin, somewhat resembling the scales in human harlequin ichthyosis caused by mutations in the ABCA12 gene.4 Transmission electron microscopy suggested the presence of lipid-like vesicles in the skin (Fig. 36.23C–D). Similar approaches have been utilized to study congenital ichthyosis due to mutations in capn12 gene and epidermal differentiation caused by mutations in svep1 gene.5,6
Further studies suggested that wound re-epithelialization occurs independently of inflammation and fibroblast growth factor signaling, essential for fibroblast recruitment and granulation tissue formation. Together, these results demonstrated that major steps and principles of cutaneous wound healing are conserved among adult mammals and adult zebrafish, making zebrafish a valuable model for studying vertebrate skin repair.
In contrast, diseases that are of late onset or slowly progressing may not be evident in the zebrafish model system. An example of such conditions is pseudoxanthoma elasticum (PXE), a slowly progressive, ectopic mineralization disorder with late onset. Injection of the abcc6a morpholino in zebrafish resulted in an early phenotype of pericardial edema and curled tail, associated with death by 8 dpf, but there was no evidence of ectopic mineralization at this stage.7 The mineralization phenotype, if developmentally corresponding to human or mouse pathogenesis of PXE, might occur later in life. In this context, the abcc6 knockdown zebrafish is not an appropriate model for PXE.
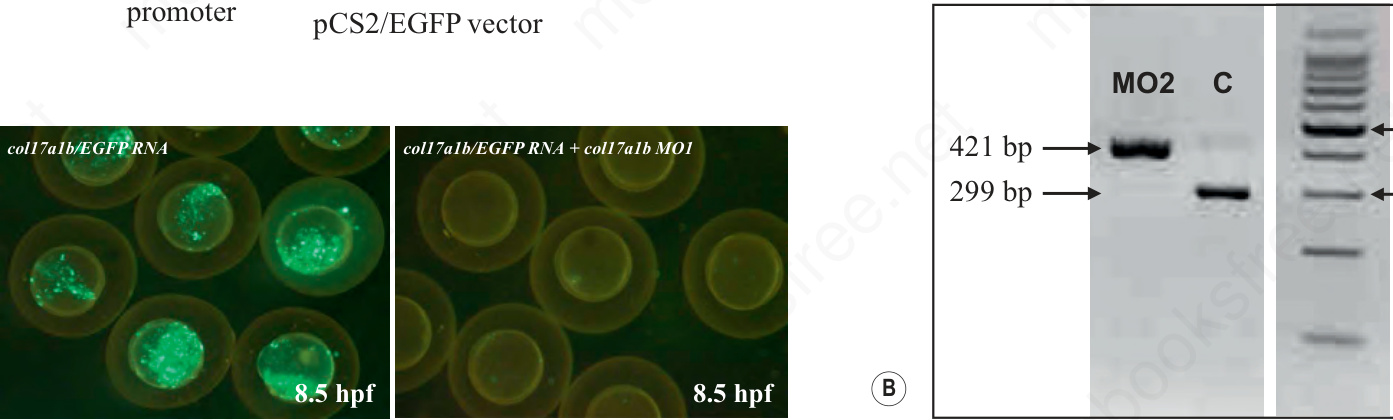
Fig. 36.22 Morpholino-mediated knockdown of zebrafish genes. (A) A morpholino (MO1) corresponding to the col17a1b gene was used to target the 5′ untranslated region of the corresponding mRNA to prevent translation. To determine the efficacy of morpholino in downregulating the translation, an expression construct consisting of SP6 promoter, 5′ UTR of the col17a1b gene, and downstream enhanced green fluorescent protein (EGFP) reporter gene was generated. Microinjection of mRNA transcribed in vitro from the pCS2/EGFP vector to 1–2 cell-stage embryo, shows green fluorescence at 8.5 hpf (lower left panel). Co-injection of this mRNA together with the MO1 morpholino completely abolished the fluorescence, indicating inhibition of the translation (bar, 1 mm). (B) A morpholino (MO2) corresponding to the zebrafish abcc6b gene was placed on the exon 18-intron 18 splice junction. Efficiency of the morpholino in preventing splicing of the abcc6b pre-mRNA into mature mRNA was monitored by RT-PCR using primers placed on exon 18 (forward) and exon 19 (reverse). PCR of the genomic sequence resulted in a 421-bp fragment, whereas fully spliced cDNA yields a 299-bp fragment devoid of intron 18 (122 bp). RT-PCR of morpholino (MO2)-treated zebrafish embryo reveals the presence of the 421-bp mRNA sequence only, indicating complete inhibition of the removal of intron 18 by splicing. As the intron 18 sequence is out of frame, this results in complete absence of the abcc6b protein product. dpf, days post fertilization; hpf, hours post fertilization; RT-PCR, reverse transcription-PCR; UTR, untranslated region. Adopted from Kim6 and Li,7 with permission.